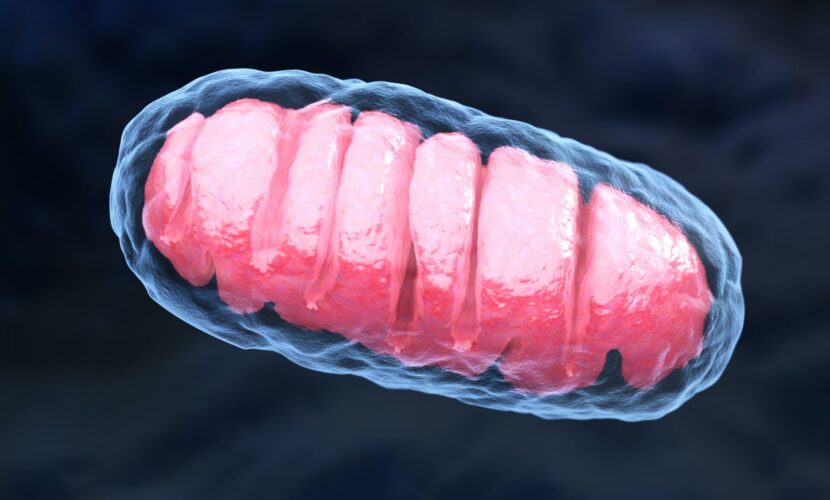
ミトコンドリアは細胞の中に存在していてエネルギーを産生しています。また、細胞内のカルシウムイオン濃度の調整や脂質の酸化、免疫反応などにも関わっています。一つの細胞に存在するミトコンドリアの数は少ない細胞で数十個、多い場合には数千個もあります。
そして、ミトコンドリア内には核とは独立したミトコンドリアゲノムが存在し二重膜構造になっています。ミトコンドリアゲノムは、αプロテオバクテリアという真正細菌に起源をもつゲノムであることが明らかになっています。
大昔に、真正細菌であったαプロテオバクテリアが古細菌の中に飲みこまれて、古細菌に飲みこまれたαプロテオバクテリアが、現在のミトコンドリアとなって真核生物に至ったと考えられています。こうして二つの細菌は共生関係に至り、膨大なエネルギーを生成する真核生物へと大きく進化しました。
ヒトのゲノムは32億塩基対にもおよぶDNA配列によって作られていますが、このうちヒト固有の遺伝子部分に相当するのはわずか1.5~2%に過ぎません。それとは対照的に、およそ50%近くのDNA配列が、過去に侵入したウイルスのDNAのなごりと考えられています。
さらにウイルスDNAのなごりが、実際にヒトの中で機能しています。その一つが女性の胎盤形成に関与するシンシチンと呼ばれるエンベロープタンパク質です。このタンパク質は、ウイルスが私たち哺乳類の祖先に侵入した際にもちこんだものとされています。
ちなみにレトロウイルスは、ヒト免疫不全ウイルス(HIV)や、ヒト成人T細胞白血病ウイルスなど逆転写酵素をもったRNAウイルスのことです。
細胞核がどのようにしてできたのかについては、細胞内共生説のような確立された学説は存在しませんでした。そうした中で細胞核ウイルス起源説が提唱されました。
ある種のウイルスは宿主の細胞内部にウイルスを生成する場を作り、それが真核生物の細胞核と非常に似ています。そのためウイルスによって作られたウイルス生成の場に、ゲノムが入ってしまった細菌が翻訳を続けられ生存できたと考えられています。
DNAは、いくつかの断片に分けられて保管されていて、アミノ酸の設計図となる塩基配列が設計図にならない塩基配列によって分断されています。アミノ酸の設計図となる部分をエキソンといい、エキソンとエキソンのあいだを分断している不要なDNA配列をイントロンと呼びます。
私たち真核生物は、DNAの配列をメッセンジャーRNAに転写する際に、このイントロンもまとめてコピーするので、それを翻訳するまでにイントロン部分を除去しなければなりません。そうした作業をスプライシングと呼びます。
そして、このイントロン・スプライシングシステムを私たち真核生物にもたらしたのも、ミトコンドリアであると考えられています。
参考文献:
・『若化!医師が見つけた若返る3つの魔法』著者 宇野克明